�z�[�� > �{�Y > �{�Y�̏�� > �L�@�Ǘ��Ő��Y�������{�Z�p��̐����w�I����э��i�ؓ���
�����E�@�{�Y�̏��@2023�N8����
�L�@�Ǘ��Ő��Y�������{�Z�p��̐����w�I����э��i�ؓ���
�k����w�@�b��w�������t�B�[���h�T�C�G���X�Z���^�[�@�u�t�@���}���@�p�B
�y�v��z
�@�ߔN�ASDGs�̊J���ڕW�ł���S�[��2�u�Q����[���Ɂv�A�S�[��13�u�C��ϓ��ɋ�̓I�ȑ���v�Ȃǂɍv�����鎝���\�Ȓ{�Y�����Ƃ��ėL�@�{�Y�ւ̎��g�݂����ڂ���Ă���B�{�����ł́A�����Ώۖq����A�L�@JAS�F���擾�����q��܂��͎擾��ڎw���q��Ƃ��A�O���X�t�F�b�h�^�C�v�A�O���C���t�F�b�h�^�C�v�Ŏ��{�������{�Z�p�틎����p���āA�e�q��̎��{�Ǘ��̏�ԁi���̂���ь��t����j�A�}�����сA���������A���i�ؓ�������͂����B
�@���̓����ł�1�������葝�̗ʂ����m�����ꂽ����A�L�@�Ǘ����n�ɂ���������̕��q�Z�p���Č�������K�v�����l����ꂽ�B�e���i�̋ؐ��ی^�\�������̓O���C���t�F�b�h�ł͋؎��k�̃G�l���M�[�Ɉˑ�����ؐ��ۂ̊����������A����͍������^�ʂ܂��͊��ԂɈˑ����邱�Ƃ����炩�ƂȂ����B�܂��A�O���X�t�F�b�h�Ő��Y���ꂽ���{�Z�p��̍��i�Ŏ��b�H�ܗL�ؐ��ۂ̊����������A���b�_�g���ł͑�ړ؋߈ʕ��ł�α���m�����_�ܗʂ����m�ȓ����ƂȂ�\������������A�O���X����уO���C���t�F�b�h�Ő��Y���ꂽ�L�@�{�Y���̊�b�I�m��������ꂽ�B
�@���̓����ł�1�������葝�̗ʂ����m�����ꂽ����A�L�@�Ǘ����n�ɂ���������̕��q�Z�p���Č�������K�v�����l����ꂽ�B�e���i�̋ؐ��ی^�\�������̓O���C���t�F�b�h�ł͋؎��k�̃G�l���M�[�Ɉˑ�����ؐ��ۂ̊����������A����͍������^�ʂ܂��͊��ԂɈˑ����邱�Ƃ����炩�ƂȂ����B�܂��A�O���X�t�F�b�h�Ő��Y���ꂽ���{�Z�p��̍��i�Ŏ��b�H�ܗL�ؐ��ۂ̊����������A���b�_�g���ł͑�ړ؋߈ʕ��ł�α���m�����_�ܗʂ����m�ȓ����ƂȂ�\������������A�O���X����уO���C���t�F�b�h�Ő��Y���ꂽ�L�@�{�Y���̊�b�I�m��������ꂽ�B
1�@�͂��߂�
�@2005�N�ɗL�@�{�Y���̕\����i�L�@JAS�i��1�j�j�����肳��A17�N���o�߂����B��Ȃǂ̔_�Y���ł͑����̔_�ꂪ�L�@JAS�F���擾���A�L�@JAS�}�[�N��t�����_�Y�����̔�����Ă���̂�ڂɂ���@��������B����A�����Ȃǂ̒{�Y���ŗL�@JAS�F���擾�������Y�s���Ǘ��҂͋ɒ[�ɏ��Ȃ��B08�N�ɒÕʗL�@���_������L�@JAS�F�؋�����̔��A09�N�ɂ͖k����w�b��w�������t�B�[���h�T�C�G���X�Z���^�[���_�q��i�ȉ��u���_�q��v�Ƃ����j�����p���ō������̗L�@JAS�F���擾�����ق��A�{���A�{���A�ؓ��ŔF�؎擾����Ă��邪�A���݂܂ŗL�@JAS�F�̒{�Y���͌���ꂽ�n��Ɛ��Y�҂Ő��Y���Ă���݂̂ŁA��ʏ���҂��w���ł���@��͂܂��܂����Ȃ��B���_�q��ł́A1994�N������q�Ǝ�������100���œ��p�����Y����̔��܂Ŏ��H�I�Ȍ������s���Ă���A�������݂̂�26�J����A���̏d880�L���O�����i��H���̏d�j�A�}���d�ʂ�527�L���O�����̃E�V�̐��Y���\�ł��邱�Ƃ��������i���}�� 2017�j�B
�@21�N5���A�_�ѐ��Y�Ȃ͎����\�ȊJ���ڕW�iSDGs�j�̒B������є_�ƕ���ɂ�����n�����g���h�~�̐���Ƃ��āu�݂ǂ�̐H���V�X�e���헪�v�����肵�A50�N�܂łɗL�@�_�Ƃ̎��g�ݖʐς��k��ʐς�25���i100���w�N�^�[���j�Ɋg�傷�邱�Ƃ�ڎw���Ă���i�_�ѐ��Y�� 2021�j�B�������Ȃ���A�L�@�{�Y�i���p���j�ɂ����Ă͗L�@JAS�F�؎擾�q�ꂪ�k�C������7�˂̂݁i22�N3�����݁j�ƂȂ��Ă���i���}�� 2022�j�A����A���y�g��Ɋւ��Ă���Ȃ���K�v�ł���B���`�Ԃ����y���Ȃ��v���́A��ʂ̗��{�Y�_�Ƃ��L�@JAS�F�؎擾�Ɋւ��葱���̃m�E�n�E��m��@����Ȃ����Ƃ�L�@�����̊m�ۂ̓���ɂ���B����ł́A17�N�ɖk�C���I�[�K�j�b�N�r�[�t�U�����c��L�@�{�Y�̕��y�g���ړI�ɐݗ�����A���Y��14�l�A���ʊW19�Ђ��������A�\���葱���A�L�@�����̊m�ہA���Y�����̗��ʂ��T�|�[�g���Ă���B�Q�悷�鐶�Y�҂̎��{�Ǘ��͂��܂��܂ŁA�O���X�t�F�b�h�i�������̂��^�j����уO���C���t�F�b�h�i�������p���^�j�A���{�Z�p���A���K�X��A���̑����G��Ǝ��{�Ǘ��ɕi���g�ݍ��킹��Ƒ���ɂ킽��B����A�L�@�{�Y�������߂����҂̓I�[�K�j�b�N�r�[�t���O���X�t�F�b�h�r�[�t�Ƃ����F���������A�M�҂͏���ґ��ւ̏�M���K�v�Ƌ���������B�܂�A����ł͗L�@JAS�F���擾�����q��̐��Y���i������������j�̓I�[�K�j�b�N�r�[�t����уI�[�K�j�b�N�z�������ƃI�[�K�j�b�N���i�Ƃ��ĂЂƂ�����ɂ���A�L�@JAS�F��L�������A���̎��{�Ǘ���i��̓��������m�ł͂Ȃ��B�]���āA�����L�@�{�Y���̉Ȋw�I�����Ɋւ�����̌������K�v�ƍl���Ă���B
�@�{�����ł͗L�@�{�Y���i���p���j���Y�̕��y�g���ړI�Ƃ��A�L�@JAS�F���擾�����A�������͎擾��ڎw���q��́A���Y�����ߒ��Ȃ�тɂ��̒{�Y���Ɋւ��鐶�Y�҂������҂܂ʼn����\�ȍ����̗L�@�{�Y���̐V���Ȏw�W�͂��邱�Ƃ�ړI�Ƃ����B
�i��1�j�u�_�ѕ����̋K�i���y�ѕi���\���̓K�����Ɋւ���@���iJAS�@�j�v�Ɋ�Â��L�@�H�i�̔F�ؐ��x�B�L�@�_�Y����L�@���H�H�i�Ȃǂ̐��Y���@�ɂ��Ă̊���߁A���̊�������̂������u�L�@�v�ƕ\���ł���悤�ɂ������̂ŁA�_�ѐ��Y�Ȃ̓o�^�F��@�ւ��F����B
�@21�N5���A�_�ѐ��Y�Ȃ͎����\�ȊJ���ڕW�iSDGs�j�̒B������є_�ƕ���ɂ�����n�����g���h�~�̐���Ƃ��āu�݂ǂ�̐H���V�X�e���헪�v�����肵�A50�N�܂łɗL�@�_�Ƃ̎��g�ݖʐς��k��ʐς�25���i100���w�N�^�[���j�Ɋg�傷�邱�Ƃ�ڎw���Ă���i�_�ѐ��Y�� 2021�j�B�������Ȃ���A�L�@�{�Y�i���p���j�ɂ����Ă͗L�@JAS�F�؎擾�q�ꂪ�k�C������7�˂̂݁i22�N3�����݁j�ƂȂ��Ă���i���}�� 2022�j�A����A���y�g��Ɋւ��Ă���Ȃ���K�v�ł���B���`�Ԃ����y���Ȃ��v���́A��ʂ̗��{�Y�_�Ƃ��L�@JAS�F�؎擾�Ɋւ��葱���̃m�E�n�E��m��@����Ȃ����Ƃ�L�@�����̊m�ۂ̓���ɂ���B����ł́A17�N�ɖk�C���I�[�K�j�b�N�r�[�t�U�����c��L�@�{�Y�̕��y�g���ړI�ɐݗ�����A���Y��14�l�A���ʊW19�Ђ��������A�\���葱���A�L�@�����̊m�ہA���Y�����̗��ʂ��T�|�[�g���Ă���B�Q�悷�鐶�Y�҂̎��{�Ǘ��͂��܂��܂ŁA�O���X�t�F�b�h�i�������̂��^�j����уO���C���t�F�b�h�i�������p���^�j�A���{�Z�p���A���K�X��A���̑����G��Ǝ��{�Ǘ��ɕi���g�ݍ��킹��Ƒ���ɂ킽��B����A�L�@�{�Y�������߂����҂̓I�[�K�j�b�N�r�[�t���O���X�t�F�b�h�r�[�t�Ƃ����F���������A�M�҂͏���ґ��ւ̏�M���K�v�Ƌ���������B�܂�A����ł͗L�@JAS�F���擾�����q��̐��Y���i������������j�̓I�[�K�j�b�N�r�[�t����уI�[�K�j�b�N�z�������ƃI�[�K�j�b�N���i�Ƃ��ĂЂƂ�����ɂ���A�L�@JAS�F��L�������A���̎��{�Ǘ���i��̓��������m�ł͂Ȃ��B�]���āA�����L�@�{�Y���̉Ȋw�I�����Ɋւ�����̌������K�v�ƍl���Ă���B
�@�{�����ł͗L�@�{�Y���i���p���j���Y�̕��y�g���ړI�Ƃ��A�L�@JAS�F���擾�����A�������͎擾��ڎw���q��́A���Y�����ߒ��Ȃ�тɂ��̒{�Y���Ɋւ��鐶�Y�҂������҂܂ʼn����\�ȍ����̗L�@�{�Y���̐V���Ȏw�W�͂��邱�Ƃ�ړI�Ƃ����B
�i��1�j�u�_�ѕ����̋K�i���y�ѕi���\���̓K�����Ɋւ���@���iJAS�@�j�v�Ɋ�Â��L�@�H�i�̔F�ؐ��x�B�L�@�_�Y����L�@���H�H�i�Ȃǂ̐��Y���@�ɂ��Ă̊���߁A���̊�������̂������u�L�@�v�ƕ\���ł���悤�ɂ������̂ŁA�_�ѐ��Y�Ȃ̓o�^�F��@�ւ��F����B
2�@�ޗ��ƕ��@
�i1�j�Ώۖq��Ƌ�������
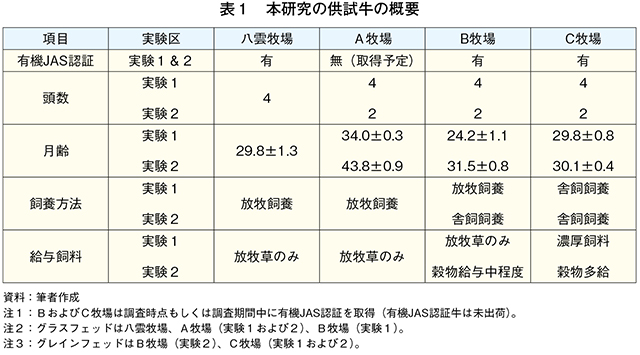
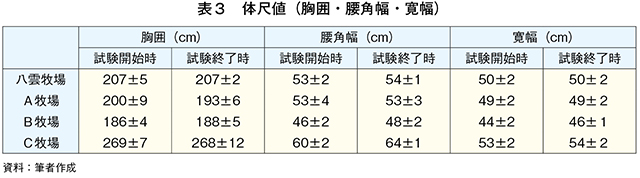
�@�{�����ɂ����鎎���Ώۖq��͗L�@JAS�F�؎擾�����A�܂��͎擾��ڎw���i�����v�擖���j�q��ł��邽�߁A���_�q��i�L�@JAS�擾�ς݁A�O���X�t�F�b�h�^�C�v�j�����A�q��i�L�@JAS�擾�\��A�O���X�t�F�b�h�^�C�v�j�AB�q��i�L�@JAS�擾�ς݁A�琬��������O���܂ł̓O���X�t�F�b�h�^�C�v�A��������̓O���C���t�F�b�h�^�C�v�j�����C�q��i�L�@JAS�擾�ς݁A�O���C���t�F�b�h�^�C�v�j�Ƃ��A���������͂��ꂼ��̖q��Ŏ��{�������{�Z�p�틎���Ƃ����i�}1�A�\1�j�B�@����1�Ƃ��āA�e�q��̎��{�Ǘ��̏�ԁi���̂���ь��t����j��c�������B���_�q��Ŏ��{�������{�Z�p�틎��4���i29.8±1.3�J����j���AA�q��Ŏ��{�������{�Z�p�틎��4���i34.0±0.3�J����j��p���A�Ċ��Ԃɕ��q�����q���ƌŌ`�������R�̐H�������BB�q��ł͓��{�Z�p�틎��4���i24.2±1.1�J����j��p���A�Ċ��Ԃɕ��q�����q�������R�̐H�Ƃ����BC�q��ł͓��{�Z�p�틎��4���i29.8±0.8�J����j��p���Ɏ����{�ō������^�Ƃ����B
�@����2�Ƃ��āA�e�q��Ő��Y���ꂽ���{�Z�p��̎}�����сA���������A���i�ؓ�������͂����B����1�ŋ����������_�q��̓��{�Z�p�틎��4���Əo�����i2021�N10���j�ƍ��킹��K�v������AA�`C�q��ł͎���1�Ƃ͈قȂ�A�܂��͂��̌Q���̋�������p�����iB�q��͏o�בO4�J���������^�j�B�e�q��A�������ɏo�ׂ��ꂽ���{�Z�p�틎���͂��ꂼ��2���iA�q��F43.8±0.9�J����AB�q��F31.5±0.8�J����AC�q��F30.1±0.4�J����j�ł������B�Ȃ��AA�q��͗L�@JAS�F�Ɍ��������q�n�ł̕��q���{�ł��邱�ƁAB�����C�q��͗L�@JAS�F���擾�ς݂ł��邪�L�@���o�ׂ܂łɎ����Ă��Ȃ����Ƃ�NjL����B

�i2�j����1 �L�@�Ǘ��Ő��Y�������p���̑��̂���ь��t����
�A �̏d����ё̎ڑ���
�@���_�q��̋������͕��q�J�n�i6���j���疈����x�A�ߑO���ɕ��s�^�̏d�v�i�L���g�����[�h�j�ŁA���̑��̖q��ł�8�������10���Ɉړ����̏d�v�i�X�}�[�g�X�P�[���j�ő̏d�𑪒肵�A�������Ԓ���1�������葝�̗ʂ��Z�o�����B�܂��A�����ɋ��́A���p���A
�C ���t����̉��
�@ ���t�����̓w�}�g�N���b�g�iHct�j�i��2�j�A�g���O���Z���h�iTG�j�i��3�j�A���R���X�e���[���iT�|CHO�j�i��4�j�A�V�����b�_�iNEFA�j�i��5�j�A�O���R�[�X�iGlucose�FGlu�j�i��6�j�R���`�]�[���i��7�j�͂����B
�i��2�j���t���ɐ�߂�Ԍ����̑̐ς̊����B
�i��3�j�������b�B
�i��4�j���t���Ɋ܂܂��R���X�e���[���i�����̈��ŁA�זE���⌌�Ǖǂ̍\���Ȃǂ̖������ʂ����j�̂��ׂĂ̗ʁB
�i��5�j���b�g�D���猌�t�ɕ��o����A�G�l���M�[���Ƃ��Ċ��p����鎉�b���B
�i��6�j���t���̃u�h�E���B�����B
�i��7�j���t�玿�z�������̈��ŒY�������A���b�A�^���p�N��ӂ𐧌䂷�����������B
�i3�j����2 �L�@�Ǘ��Ő��Y�������{�Z�p��̎}�����тƓ�������э��i�ؓ���
�A �}������
�@���ׂĂ̋�������2021�N10�����܂łɏo�ׂ���A���v�Вc�@�l���{�H���i�t����̔��s���鋍�}���i�t��������Ɏ}�����т���͂����B
�C �����g�D�̏d�ʑ���Ƒ��ݓ����̉�U�w�I���
�@�d�ʑ����̑��݂���U�w�I�ɕ��ނ���镬�啔�A���E����A�ؒ��ߕ��A�w
�E ���i�̑g�D���w�I���
�G ���b�_�g���Ɖh�{����
�@���͂ɗp���鋹�Œ�����ё�ړ؋߈ʕ��͔牺���b����菜���A�\�Ȍ���Ԑg�����݂̂��~���`��ɂ��A���͂܂Ő^���ԁE�}�C�i�X30�x�ŕۑ������B�@���b�_�g���̑���̓~���X�`���_�iC14�F0�j�A�p���~�`���_�iC16�F0�j�A�p���~�g�I���C���_�iC16�F1�j�A�X�e�A�����_�iC18�F0�j�A�I���C���_�iC18�F1�j�AC18�F1���b�_�ِ̈��́AC18���b�_�ِ̈��́A���m�[���_�iC18�F2�j�����α�|���m�����_�iC18�F3�j�̎��b�_���`���G�X�e���̊ܗL�����Z�o�����B
�@�h�{�������͔͂M�ʁA�����A�^���p�N���A�����A�Y�������ɂ��Ĉ�ʍ��c�@�l�H�i���͊J���Z���^�[SUNATEC�Ɉϑ����A���肵���B
�@����1�����2�ʼn�͂��ꂽ���ڂ���L�@�{�Y�����H����q��̉Ȋw�I���ʓ_�𒊏o���A�L�@JAS�F�؎擾�q��Ő��Y���ꂽ�{�Y���i���Y�ߒ����܂ށj�ɂ�����Ȋw�I�w�W�͂����B
3�@���ʂƍl�@
�i1�j�̏d����ё̎ڑ���
�@�\2�ɋ������̌���A�����J�n���i2021�N8���j�̑̏d�A�����I�����i21�N10���j�̑̏d�A�������Ԓ���1�������葝�̗ʂ��A�\3�ɋ��́A���p���A�����������B�����J�n������яI�����̑̏d�̈Ⴂ�͌���Ƃ��ꂼ��̎��{�Ǘ��ɋN������B�̍ގ��̎��{�Ǘ��́A���_�q��AA�q��AB�q�ꂪ���q���{�ł���A���_�q�ꂪ�L�@�Ǘ��Œʏ�q���^�i����킪�y���j�A�����C�O���X�A�I�[�`���[�h�O���X�A�V���N���[�o�[�j�AA�q�ꂪ�L�@�Ǘ��̖쑐�^�i����킪�m�R���M�N�A�J�Y�U�L�����M�j�AB�q�ꂪ�L�@�Ǘ��̒ʏ�q���^�i����킪�I�[�`���[�h�O���X�A���[�h�J�i���[�O���X�j�̊��S�O���X�t�F�b�h�ł���B����AC�q��͎Ɏ����{�ŃR�[���T�C���[�W�A�r�[�g�p���v�A�K�i�O�����A�����Ȃǂ�TMR�iTDN70���ȏ�j�̔z�����������^����O���C���t�F�b�h�ł���B�]���āA�̍��͑傫����C�q��̐��̏d�����1�������葝�̗ʂ����q���荂�����Ƃ͎Ɏ����{�ɂ��s���ʂ̗}���Ƌ��^�����ɂ��e���̉\�����l����ꂽ�B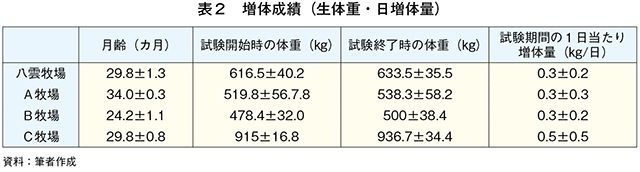
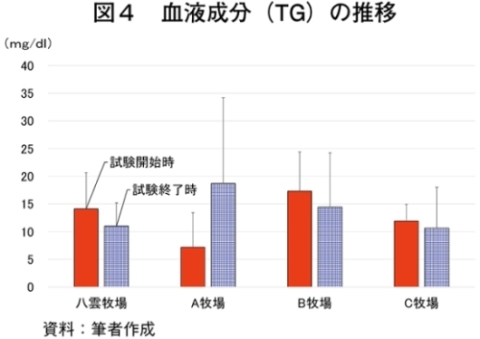
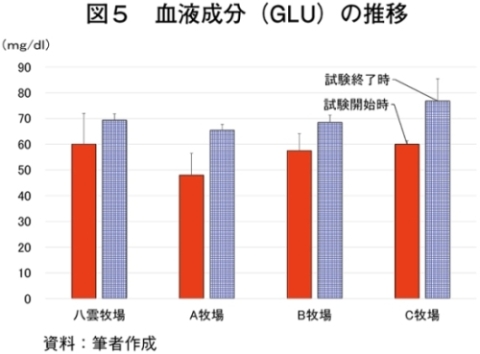
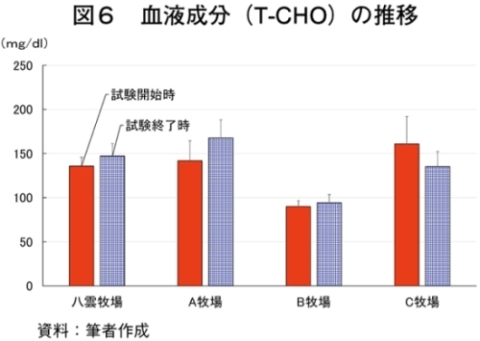
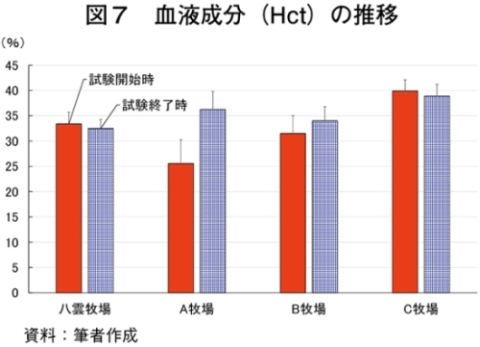
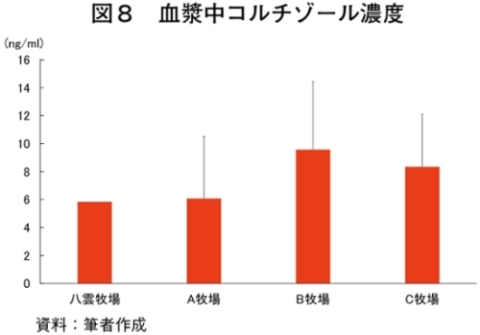
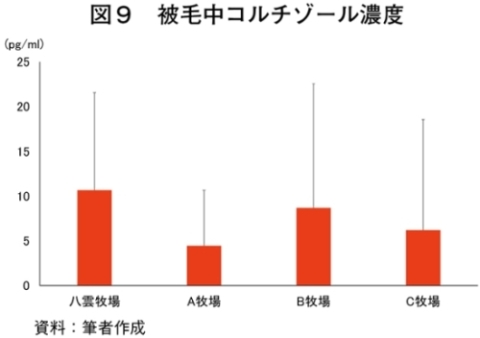
�i2�j��ʌ��t�����̐���
�@�������̌��N��Ԃ�c�����邽�߂Ɉ�ʌ��t�����̐��ڂ���͂����Ƃ���A����TG�Z�x�������i���ۂ� 2014�j�A���s���̊�l�͈͓̔��iNEFA�F0�`0.4 mEq/ml�i�}3�j�ATG�F30�`70mg/dl�i�}4�j�AGLU�F61.5�`69.5mg/dl�i�}5�j�AT-CHO�F99.8�`120.8 mg/dl�i�}6�j�AHct�F27�`34���i�}7�j�j�ł������B����NEFA�Z�x�͑̎��b�����̎w�W�Ƃ���ATG�Z�x�͒������b�̎w�W�Ƃ��Ď����s���Œቺ�AT-CHO�Z�x�̓G�l���M�[�[���̎w�W�AGLU�Z�x�̓G�l���M�[�s����ԂŒቺ����i�{�Y�Z�p���� 2005�j�B��l�͈͊O�ł���������TG�Z�x�͎��b���G�d���̋����Y�����ŏd�v�Ȉ��q�ł��邪�A�O���X�t�F�b�h����уO���C���t�F�b�h�ŗL�ӂȍ��͂Ȃ��A�O���X�t�F�b�h�ł͉ߓx�Ȏ��b���G��K�v�Ƃ��Ȃ��Ԑg�������Y��ړI�Ƃ��Ă��邽�߁A����TG�Z�x�̒ቺ�i��l�O�j�͕��q�Ǝ����e�����݂̂œ��p���Y����ꍇ�A�l������K�v���Ȃ��\�������������B�܂��A�O���C���t�F�b�h�ł͌���TG�Z�x���T-CHO�����GLU�AHct�Z�x���O���X�t�F�b�h��茰���ɍ����A���^�����̉e�������f���ꂽ�B�������Ȃ���A�����̃f�[�^�͒Z���I�ȉ�͂ł���A��ʌ��t�����̐��ڂ��p���I�ɍs���K�v������B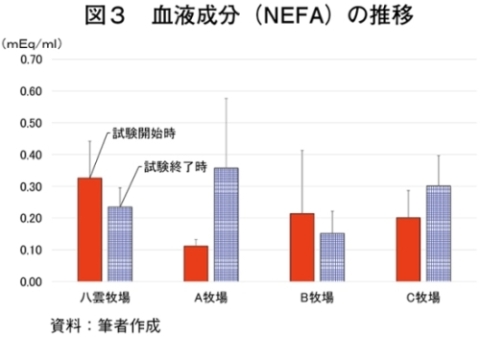
�@�L�@�{�Y�̓A�j�}���E�F���t�F�A�ɔz���������Y�����ł��邽�߁i�_�ѐ��Y�� 2021�j�A�X�g���X�w�W�ŗp������R���`�]�[���������I�����̌�
�i3�j�}������
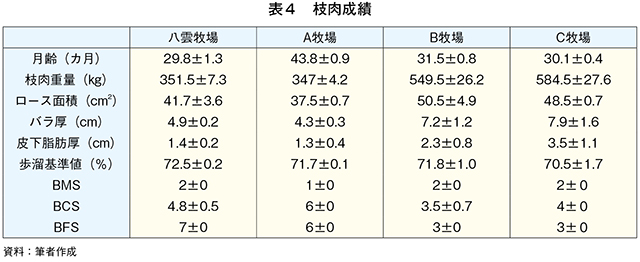
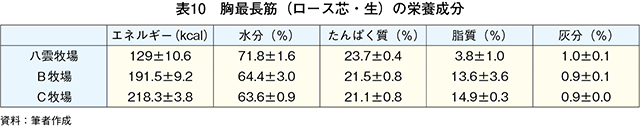
�@�O���X�t�F�b�h�i���_�q�ꂨ���A�q��j����уO���C���t�F�b�h�iB�q�ꂨ���C�q��j�̓��{�Z�p��̎}�����т���͂����i�}10�A�\4�j�BB�q��ł͏o��4�`6�J���O��荒�����^���s�����߁A�O���X�t�F�b�h����Ɏ����{�̃O���C���t�F�b�h�ɐ�ւ��B�O���X�t�F�b�h�ł̓O���C���t�F�b�h���}���d�ʁA���[�X�ʐρA�o�����A�牺���b�����Ⴍ�ABCS�i�����F��j�����BFS�i�����b�F��j�����������B�}���d�ʁA���[�X�ʐρA�o�����A�牺���b�����Ⴂ���Ƃ͐ێ�G�l���M�[�ƕ��q���{�ɂ��^���s���i�G�l���M�[����j�̍��ɋN������ƍl����ꂽ�BBCS�ł͕��q�s���ő�������~�I�O���r���̑����i�ɓ��� 1989�j�ABFS�ł͐ێ掔���ł�����q���R����β�J���`���̒����ɋN������i������ 2010�j�B���b���G��ł���BMS�i�����b���G��j�Ɋւ��Ă̓O���C���t�F�b�h�ƃO���X�t�F�b�h�ɖ��m�ȍ��͂Ȃ��A���{�Z�p��̎��b���G�\�͂̒Ⴓ�������ꂽ�B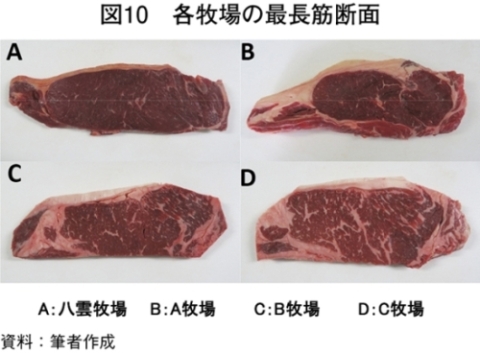
�i4�j�����g�D�̏d�ʂƑ��ݓ����̉�U�w�I���
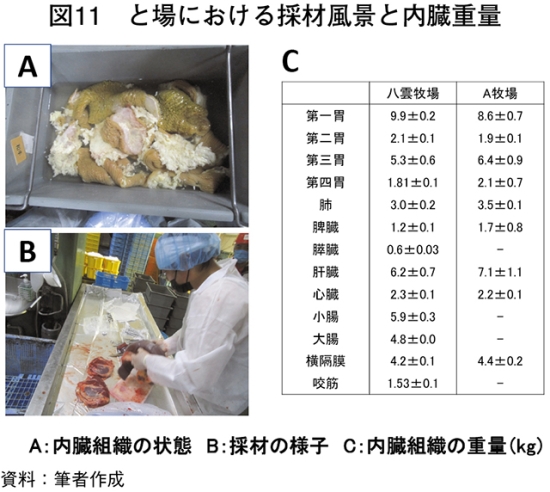
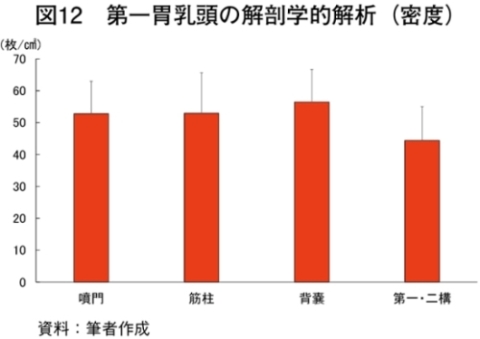
�@�O���X�t�F�b�h�̓����g�D�̏d�ʂ�}11�Ɏ����BB�����C�q��̋������̓����g�D�͔p���܂��͐��Y�҂���ї��ʋƎ҂���̋����������Ȃ��������߁A�̍ނł��Ȃ������B�@���s�̒{�Y�����ł͚M�����Ԃ͒����Ă�3�J����ł������A�L�@�{�Y���H�q��ł͚M�����Ԃ̒�������������i���_�q�ꂨ���A�q���6�J����܂ŚM�����ԁj�B�M�����Ԃ̒������͑��ݓ����i�E�V�̐��̃G�l���M�[�A�Y�����A�Y�����Ɋ֗^������������b�_���z������j�̔��B���x������B���ݓ����̔��B�ɂ͑e�����Ȃǂ̕����I�h������э��������R���̃v���s�I���_�A���_�ɂ�鉻�w�I�h�����K�v�Ƃ���邽�߂ł���i�Óc 1963�j�B�{�����ł͔��_�q��Ő��Y�������{�Z�p��̑��ݓ����ɂ����āA���x�A���a�A�Z�a�A�\�ʐς���͂����Ƃ���A�e���ʂŗL�ӂȍ��͔F�߂��Ȃ������i�}12�`15�j�B���s���̃E�V�ł͑��ݓ����̖��x�͕���A�ؒ��A�w�X�A���E��\�ł��ꂼ��A38.5�A47.5�A33.5�A47.0��/�����Z���`���[�g���A���a�̕��ϒl�ł�0.5�A0.2�A0.1�A0.4�Z���`���[�g���ł���i�Óc 1966�j�A�{���Ƃ̃O���X�t�F�b�h�Ő��Y���ꂽ���{�Z�p��͍������^�Ɠ����ȏ�̑��ݓ����̔��B��L���A�e�����ɂ�镨���I�h���ŏ\���ɑ��ݓ����̔��B��������邱�Ƃ����炩�ƂȂ����B
�i5�j���i�̑g�D���w�I���
�@�ؐ��ی^�\�������͉ƒ{����щƂ�����ɂ����āA�H��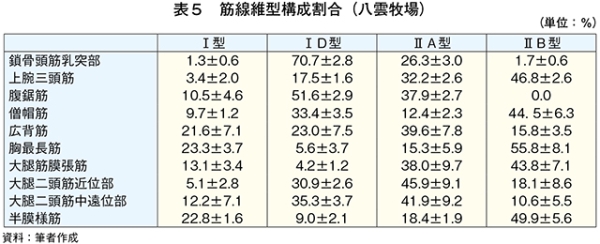
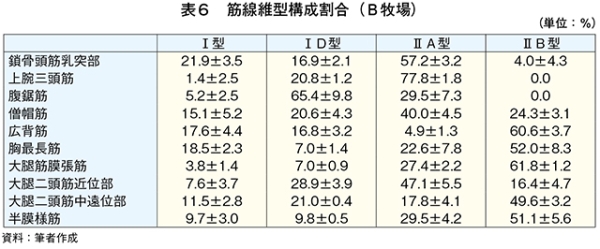
�@�T����чTD�^�ؐ��ۂ͎�Ƃ��Ď��b�_���؎��k�G�l���M�[�Ƃ��ė��p����B���q���{�ł͍s���ʂ��������邱�ƁA�O���X�t�F�b�h�ł͔牺���b���������X���ɂ��邱�Ƃ�����̓��̎��b�����ċؑg�D�ɃG�l���M�[���Ƃ��ē�������A�TD�^�ؐ��ۂ���������Ɛ��������i���}�� 2017�j�B
�@����A�O���C���t�F�b�h�ɂ����Ă����ʂ̇TD�^�ؐ��ۂ����݂��鍜�i�����݂����B�ؐ��ی^�̈ڍs�͉^�����ׂ������͐ێ掔���̈Ⴂ���琶���A�����ܗʂ̍���������ێ悵���ꍇ�A�x�،^�A�܂�T�^����чTD�^�ؐ��ۂɈڍs����iWang et al., 2013�j�iMaxwell et al.,2014�j�B�O���C���t�F�b�h�ŔF�߂�ꂽ�TD�^�ؐ��ۂ̑��݂͐ێ掔���̉e���ɂ��\�����������ꂽ�B
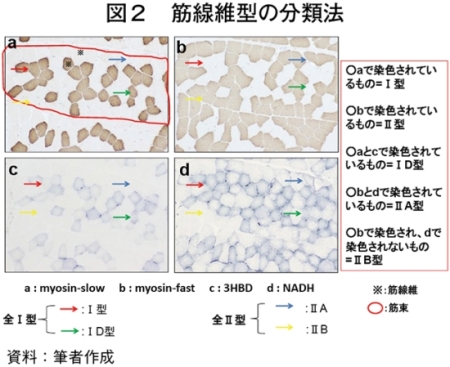
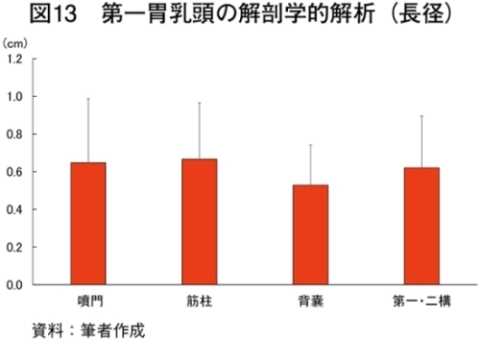
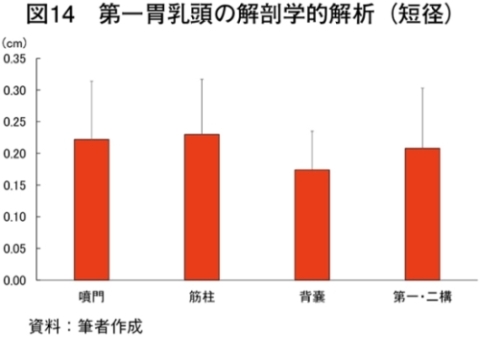
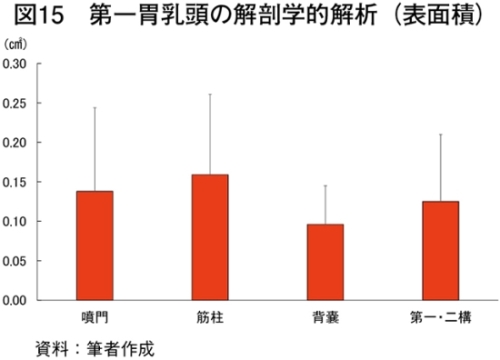
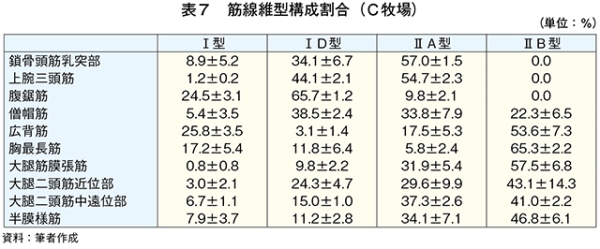
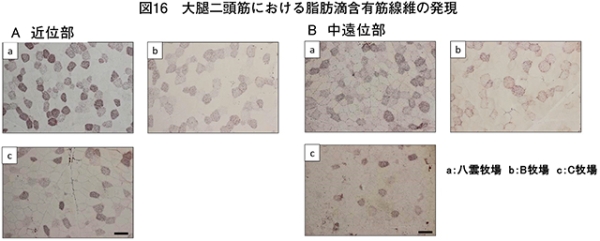
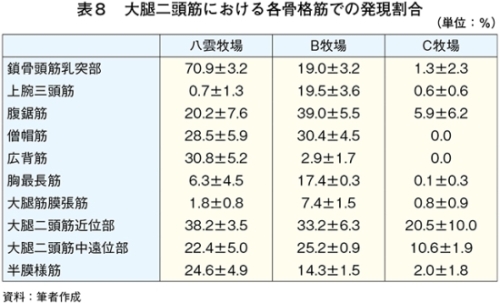
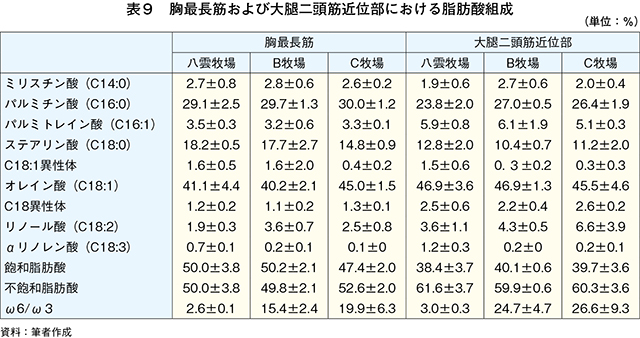
�@�}16����ѕ\8�Ɋe�q��̓��{�Z�p��̑�ړ؋߈ʕ�����ђ����ʕ��ɂ����鎉�b�H�i��8�j�ܗL�ؐ��ۂ̐��F���Ƃ��̔��������������B
�@���b�H�ܗL�ؐ��ۂ̑����͋ؐ��ی^�ڍs�Ɠ��l�ɉh�{�ێ您��щ^�����ׂɂ�萶����\������������邪�A��
�i��8�j�זE���ɑ��݂���A������^���p�N���Ȃǂ��܂ދ��`�̉t�̂̌ł܂�B
�i6�j���b�_�g���Ɖh�{����
�@���b�H�ܗL�ؐ��ۂ̊����������ɈقȂ������ʂł��鋹�Œ�����ё�ړ؋߈ʕ��̎��b�_�g����\9�Ɏ����B��ʓI�ɋ����̓I���C���_�̊����������A�p���~�`���_�ƃX�e�A�����_�̊��������Ȃ��قǕ������ǂ��Ƃ���A�I���C���_�̊����ƕ����̍D�܂����ɐ��̑��ւ����邱�Ƃ�����Ă���iMandall et al., 1998�j�i���{ 2012�j�B���Œ��ɂ����Ă̓O���C���t�F�b�h�ŃI���C���_�����������B����A��ړ؋߈ʕ��ł�α���m�����_�ȊO�̎��b�_�ō��͔F�߂��Ȃ������Bα���m�����_�͖q���R���̎��b�_�ł��邱�Ƃ���O���X�t�F�b�h�̎��{�Ǘ��ő������邱�Ƃ͐����ł���Bα���m�����_�̓q�g�̑̓��ł͐��Y�ł��Ȃ��K�{���b�_�Ō������P�⓮���d���̗\�h�ɓ����i�N�� 2007�j�B��ړ؋߈ʕ��ł͎��b�H�ܗL�ؐ��ۂ����{�Ǘ��Ɋւ�炸���������ő��݂��A�����α���m�����_���������������邽�߁A��ړ؋߈ʕ��ł�α���m�����_�ܗʂ��O���X�t�F�b�h�^�L�@�{�Y�Ő��Y�������{�Z�p��̖��m�ȓ����ƂȂ�\�����������ꂽ�B�@���Œ��̉h�{�����i�M�ʁA�����A�^���p�N���A�����A�D���j��\10�Ɏ����B�O���X�t�F�b�h�ł͐����A�^���p�N���������A�G�l���M�[�A�������Ⴉ�����B���̌X���͍������^�ʂ���ы��^���ԂɈˑ������iC�q�ꁃB�q�ꁃ���_�q��j�B
4�@������
�@�{�����ł͎����Ώۖq���L�@JAS�F�؎擾�����q��܂��͎擾��ڎw���i�����v�擖���j�q��Ƃ��A�O���X�t�F�b�h�^�C�v�A�O���C���t�F�b�h�^�C�v�Ŏ��{�������{�Z�p�틎����p���āA����1�Ƃ��Ċe�q��̎��{�Ǘ��̏�ԁi���̂���ь��t����j����͂����B���Ɏ���2�Ƃ��Ċe�q��Ő��Y���ꂽ���{�Z�p��̎}�����сA���������A���i�ؓ�������͂����B���̓����ł͕��q���{���̔����̓��{�Z�p���1�������葝�̗ʂ����m�����A�O���X�t�F�b�h�^�L�@�{�Y�Ő��Y������{�Z�p��̑��̗ʂ������邽�߂ɂ́A�L�@�Ǘ����n�ɂ���������̕��q�Z�p�i�h�{���̍�������\���Ȃǁj���Č�������K�v�����l����ꂽ�B����ŁA�O���X�t�F�b�h�^�L�@�{�Y�ł͔�����1�������葝�̗ʂ͖�0.3�L���O�����ł��������Ƃ��獒�������̊��s���{�Ɣ�r���Ĉ��̑��̂����߂�ꍇ�A�o���Ԃ�����������\�����l����ꂽ�B���ꂪ���Y�o��ɒ��ړI�ɉe�����A��ʎs��ŗ��ʂ���L�@�{�Y���������ɂȂ蓾�����ɂȂ���ƍl������B���t�����̐��ڂł͂��ׂĂ̎��{�Ǘ��Ō���TG�Z�x�������A���s���̊�l�͈͓̔��ł��邱�Ƃ����炩�ƂȂ����B���݂̉�U�w�I��͂ł͑e�����݂̂ŏ\���ȑ��ݓ����̔��B��������邱�Ƃ����炩�ƂȂ�A�O���X�t�F�b�h�Ő��Y�������{�Z�p��̑��݂̉�U�w�I�m��������ꂽ�B�e���i�̋ؐ��ی^�\�������̓O���C���t�F�b�h�ł͋؎��k�̃G�l���M�[�Ɉˑ����鑬�،^�̇U�^�i�UA�^�{�UB�^�j�ؐ��ۂ̊����������A����͍������^�ʂ܂��͊��ԂɈˑ����邱�ƁA�O���C���t�F�b�h�ɂ����Ă����ʂ̇TD�^�ؐ��ۂ����݂��鍜�i�����݂��邱�Ƃ����炩�ƂȂ����B�܂��A�O���C���t�F�b�h���O���X�t�F�b�h�Ő��Y���ꂽ���{�Z�p��̍��i�Ŏ��b�H�ܗL�ؐ��ۂ̊����������A���b�_�g���ł͑�ړ؋߈ʕ��ł�α���m�����_�ܗʂ��L�@�Ǘ��Ő��Y�����O���X�t�F�b�h���{�Z�p��̖��m�ȓ����ƂȂ�\�����������ꂽ�B
�@�ȏ���A�L�@�Ǘ����Ő��Y�����O���X�t�F�b�h����уO���C���t�F�b�h�̓��{�Z�p��̓����͊e���ڂŖ��炩�ƂȂ�A�{�����ɂ��M�����Ԃ�����������\���̍����O���X�t�F�b�h�^�L�@�{�Y�ɂ����āA���ݓ����̔��B�͍������^���Ɠ����ȏ�ł��邱�ƁA�����{�Ǘ��ł̊e���i�̋ؐ��ی^�\�������Ǝ��b�_�g���̊�b�I���i�L�@�{�Y���ɂ����Ă������ܗʂ������_�炩�����������߂����҂̓O���C���t�F�b�h���A�����ܗʂ��Ⴂ�Ԑg���������߂����҂̓O���X�t�F�b�h���j�������ꂽ�B�������Ȃ���A�����{�Ǘ��ŋ��ʂ�������͌����������A����A�L�@�{�Y���Ɋւ��邳��Ȃ�f�[�^�̒~�ς��]�܂��B�Ō�ɖ{���Ƃ̐��ʂ����̐�A�L�����������L�@�{�Y�̔��W�Ƃ���ɔ����L�@�{�Y�̐��Y�҂���я���ґo���ւ̊�b�I������邱�Ƃɂ�镁�y�g��A���Ȃ킿�����ɂ����鎝���\�Ȓ{�Y�̌n�̍\�z�̈ꏕ�ɂȂ��邱�Ƃ����҂���B
�� ��
�{�����́A�e�q��A�k����w�b��w���h�{�����w�������̗����D�y�����A�}���n�j�`��������Ђ̊����ۉے��㗝�̂����͂����ƂŐ��s�ł��܂����B�����Ɋ��ӂ̈ӂ������܂��B
�Q�l����
1�j�ɓ��ǁC�L���\�O�C�ߓ��m�C�M���a�K�C�{���T�C�r�c�o�D�z���X�^�C���틎�����}���̋����F��ɉe�����鏔�v���̉�́D���{�{�Y�w��C1990�C61�i6�j�C520�|526�D
2�j���}���p�B�D�k�����_���`�������Ő��Y�E�̔��E�������鋍���`�D�H���̉Ȋw�C2017�C151�\157�D
3�j���}���p�B�D���_�q�ꂪ���H���鎝���\�Ȓ{�Y�\�k�����_���̐��Y����̔��A���y�܂Ł[�D�{���̗F�C2022�C49�|53�D
4�j���ۍF�V��C�u��p�b�C���C�~�q�C���c�F�i�D���������ɂ����鍕�јa���狎�����̌��t�����w�I����D�Y�Ɠ����Տ���w�G���C2015�C5�i4�j�C185�\190�D
5�j�N��q���C�|���O�K�C�֍����j�C���쌫���C�R�q���C�ߓ��a�Y�Dα�\���m�����_�ێ�ɂ�錌���ቺ��p�Ɋւ��錟���D���{���a�V�X�e���w��G���D2007�C13�i2�j�C331�\333�D
6�j�Вc�@�l�{�Y�Z�p����D�ƒ{�q���}�j���A���C2005�Dhttp://jlta.lin.gr.jp/report/detail_oversea_pdf/kaigai_m036.pdf
7�j��ؗR���q�C���}���p�B�C�~���a�O�C�c������C�����ˉ�V�D�L�@�I�Ǘ����H�q��ɂ�����A���Ƌ��̍̐H�s���̊W�C�_�ƐH���H�w��k�x����C2014�C��61���ʍ��D
8�j�Óc�P�V�D���݂̐����D���{�{�Y�w���C1963�C34�i4�j�C235�\242�D
9�j�Óc�P�V�D�����̉Ȋw�C1966�D
10�j�����D���C���쐴�C�����Y��A�_�J�[�C��Ήh��C���������C�ы`�N�C�R�c�����D���q�Ǝ���������g�ݍ��킹���V�������p�����Y���@“���n�琬�E���Z�p”�̉\���Ɠ����̓����C���{�g�n�{�Y�w���C2010�C53�i1�j�C41�|49�D
11�j�_�ѐ��Y�ȁD�݂ǂ�̐H���V�X�e���헪�C2021�Dhttps://www.maff.go.jp/j/kanbo/kankyo/seisaku/midori/attach/pdf/index-10.pdf
12�j�_�ѐ��Y�ȁD�L�@�{�Y���̐��Y�H���Ǘ��҃n���h�u�b�N�C2021�Dhttps://www.maff.go.jp/j/jas/jas_kikaku/attach/pdf/yuuki-302.pdf
13�j�щp���D�V���Ȑ����w�I�X�g���X�]���@�Ƃ��̉��p�D2018�C62�i2�j�C29�\32�D
14�j���{�a�T�D�ŐV�_�ƋZ�p �{�Yvol�D4�i�_��������������ҁj�C2012�C76�D
15�j�n粍N��D�ؐ��ی^�ƐH���̂��������\��䍓������ٓI�T�^�ؐ��ۂ̎Y���_�I�@�\�`�Ԋw��́\�D���k�{�Y�w���C2016�C66�C1�\6�D
16�jGap-Don Kim, Youn-Chul Ryu�CJin-Yeon Jeong�CHan-Sul Yang�CSeon-Tea Joo�DRelationship between pork quality and characteristics of muscle fibers classified by the distribution of myosin heavy chain isoforms�C Journal of Animal Science�D2013�C91�C5525�|5534�D
17�jHideaki Hayashi�CChigusa Arai�CYurie Ikeuchi�CMaho Yamanaka�DEffect of growth and parturition on hair cortisol in Holstein cattle�DAnimal Science Journal�C2021�C92�i1�j�D
18�jDaisuke Mashima�CYoshiaki Oka�CTakafumi Gotoh�CShozo Tomonaga�CShoko Sawano�CMako Nakamura�CRyuichi Tatsumi�CWataru Mizunoya�DCorrelation between skeletal muscle fiber type and free amino acid levels in Japanese Black steers�DAnimal Science Journal�C2019�C90�C604�\609�D
19�jMandall IB�CJ.G.Buchanan-Smith�CC.P.Campbell�DEffects of forage vs grain feeding on carcass characteristics, fatty acid composition, and beef quality in Limousin-cross steers when time on feed is controlled�D Journal of Animal Science�C1998�C76�C2619�\2630�D
20�jMaxwell S. DeNies�CJordan Johnson�CAmanda B. Maliphol�CMichael Bruno�CAnnabelle Kim�CAbbas Rizvi�CKevyn Rustici�CScott Medler�DDiet-induced obesity alters skeletal muscle fiber types of male but not female mice�CPhysiological Reports�C2014�C2�i1�j�C204�D
21�jYichen Wang�CJeffrey E. Pessin�DMechanisms for fiber-type specificity of skeletal muscle atrophy�CCurrent Opinion in Clinical Nutrition and Metabolic Care�C2013�C16�i3�j�C243�\250�D
�@�ȏ���A�L�@�Ǘ����Ő��Y�����O���X�t�F�b�h����уO���C���t�F�b�h�̓��{�Z�p��̓����͊e���ڂŖ��炩�ƂȂ�A�{�����ɂ��M�����Ԃ�����������\���̍����O���X�t�F�b�h�^�L�@�{�Y�ɂ����āA���ݓ����̔��B�͍������^���Ɠ����ȏ�ł��邱�ƁA�����{�Ǘ��ł̊e���i�̋ؐ��ی^�\�������Ǝ��b�_�g���̊�b�I���i�L�@�{�Y���ɂ����Ă������ܗʂ������_�炩�����������߂����҂̓O���C���t�F�b�h���A�����ܗʂ��Ⴂ�Ԑg���������߂����҂̓O���X�t�F�b�h���j�������ꂽ�B�������Ȃ���A�����{�Ǘ��ŋ��ʂ�������͌����������A����A�L�@�{�Y���Ɋւ��邳��Ȃ�f�[�^�̒~�ς��]�܂��B�Ō�ɖ{���Ƃ̐��ʂ����̐�A�L�����������L�@�{�Y�̔��W�Ƃ���ɔ����L�@�{�Y�̐��Y�҂���я���ґo���ւ̊�b�I������邱�Ƃɂ�镁�y�g��A���Ȃ킿�����ɂ����鎝���\�Ȓ{�Y�̌n�̍\�z�̈ꏕ�ɂȂ��邱�Ƃ����҂���B
�� ��
�{�����́A�e�q��A�k����w�b��w���h�{�����w�������̗����D�y�����A�}���n�j�`��������Ђ̊����ۉے��㗝�̂����͂����ƂŐ��s�ł��܂����B�����Ɋ��ӂ̈ӂ������܂��B
�Q�l����
1�j�ɓ��ǁC�L���\�O�C�ߓ��m�C�M���a�K�C�{���T�C�r�c�o�D�z���X�^�C���틎�����}���̋����F��ɉe�����鏔�v���̉�́D���{�{�Y�w��C1990�C61�i6�j�C520�|526�D
2�j���}���p�B�D�k�����_���`�������Ő��Y�E�̔��E�������鋍���`�D�H���̉Ȋw�C2017�C151�\157�D
3�j���}���p�B�D���_�q�ꂪ���H���鎝���\�Ȓ{�Y�\�k�����_���̐��Y����̔��A���y�܂Ł[�D�{���̗F�C2022�C49�|53�D
4�j���ۍF�V��C�u��p�b�C���C�~�q�C���c�F�i�D���������ɂ����鍕�јa���狎�����̌��t�����w�I����D�Y�Ɠ����Տ���w�G���C2015�C5�i4�j�C185�\190�D
5�j�N��q���C�|���O�K�C�֍����j�C���쌫���C�R�q���C�ߓ��a�Y�Dα�\���m�����_�ێ�ɂ�錌���ቺ��p�Ɋւ��錟���D���{���a�V�X�e���w��G���D2007�C13�i2�j�C331�\333�D
6�j�Вc�@�l�{�Y�Z�p����D�ƒ{�q���}�j���A���C2005�Dhttp://jlta.lin.gr.jp/report/detail_oversea_pdf/kaigai_m036.pdf
7�j��ؗR���q�C���}���p�B�C�~���a�O�C�c������C�����ˉ�V�D�L�@�I�Ǘ����H�q��ɂ�����A���Ƌ��̍̐H�s���̊W�C�_�ƐH���H�w��k�x����C2014�C��61���ʍ��D
8�j�Óc�P�V�D���݂̐����D���{�{�Y�w���C1963�C34�i4�j�C235�\242�D
9�j�Óc�P�V�D�����̉Ȋw�C1966�D
10�j�����D���C���쐴�C�����Y��A�_�J�[�C��Ήh��C���������C�ы`�N�C�R�c�����D���q�Ǝ���������g�ݍ��킹���V�������p�����Y���@“���n�琬�E���Z�p”�̉\���Ɠ����̓����C���{�g�n�{�Y�w���C2010�C53�i1�j�C41�|49�D
11�j�_�ѐ��Y�ȁD�݂ǂ�̐H���V�X�e���헪�C2021�Dhttps://www.maff.go.jp/j/kanbo/kankyo/seisaku/midori/attach/pdf/index-10.pdf
12�j�_�ѐ��Y�ȁD�L�@�{�Y���̐��Y�H���Ǘ��҃n���h�u�b�N�C2021�Dhttps://www.maff.go.jp/j/jas/jas_kikaku/attach/pdf/yuuki-302.pdf
13�j�щp���D�V���Ȑ����w�I�X�g���X�]���@�Ƃ��̉��p�D2018�C62�i2�j�C29�\32�D
14�j���{�a�T�D�ŐV�_�ƋZ�p �{�Yvol�D4�i�_��������������ҁj�C2012�C76�D
15�j�n粍N��D�ؐ��ی^�ƐH���̂��������\��䍓������ٓI�T�^�ؐ��ۂ̎Y���_�I�@�\�`�Ԋw��́\�D���k�{�Y�w���C2016�C66�C1�\6�D
16�jGap-Don Kim, Youn-Chul Ryu�CJin-Yeon Jeong�CHan-Sul Yang�CSeon-Tea Joo�DRelationship between pork quality and characteristics of muscle fibers classified by the distribution of myosin heavy chain isoforms�C Journal of Animal Science�D2013�C91�C5525�|5534�D
17�jHideaki Hayashi�CChigusa Arai�CYurie Ikeuchi�CMaho Yamanaka�DEffect of growth and parturition on hair cortisol in Holstein cattle�DAnimal Science Journal�C2021�C92�i1�j�D
18�jDaisuke Mashima�CYoshiaki Oka�CTakafumi Gotoh�CShozo Tomonaga�CShoko Sawano�CMako Nakamura�CRyuichi Tatsumi�CWataru Mizunoya�DCorrelation between skeletal muscle fiber type and free amino acid levels in Japanese Black steers�DAnimal Science Journal�C2019�C90�C604�\609�D
19�jMandall IB�CJ.G.Buchanan-Smith�CC.P.Campbell�DEffects of forage vs grain feeding on carcass characteristics, fatty acid composition, and beef quality in Limousin-cross steers when time on feed is controlled�D Journal of Animal Science�C1998�C76�C2619�\2630�D
20�jMaxwell S. DeNies�CJordan Johnson�CAmanda B. Maliphol�CMichael Bruno�CAnnabelle Kim�CAbbas Rizvi�CKevyn Rustici�CScott Medler�DDiet-induced obesity alters skeletal muscle fiber types of male but not female mice�CPhysiological Reports�C2014�C2�i1�j�C204�D
21�jYichen Wang�CJeffrey E. Pessin�DMechanisms for fiber-type specificity of skeletal muscle atrophy�CCurrent Opinion in Clinical Nutrition and Metabolic Care�C2013�C16�i3�j�C243�\250�D