ホーム > 畜産 > 畜産の情報 > ヒトの匂い知覚メカニズムを応用する牛肉香の“みえる化”
調査・報告 畜産の情報 2023年9月号
ヒトの匂い知覚メカニズムを応用する牛肉香の“みえる化”
静岡県立大学 食品栄養科学部 准教授 伊藤 圭祐
【要約】
畜肉において、匂いはおいしさを決定づける非常に重要な要素である。ヒトは約400種類の嗅覚受容体の応答パターンによって1兆種類以上の匂いを感知・識別している。本研究では、ヒト嗅覚受容体の応答パターンを網羅的に解析することで、2種類の牛肉(米国産牛肉と黒毛和牛肉)の匂いの“みえる化”を試みた。その結果、各牛肉は特有のヒト嗅覚受容体応答パターンを示し、米国産牛肉ではhOR6C6、hOR52K1、hOR1M1など、黒毛和牛肉ではhOR4F6、hOR4F16、hOR13F1などが応答受容体として検出された。従来行われてきた官能評価や成分分析に加え、ヒト嗅覚受容体応答を指標とする新しいアプローチを組み合わせることで、畜肉のおいしさの客観的・定量的な評価が可能になると期待できる。
1 はじめに
(1)畜肉のフレーバー形成について
食品のおいしさには味に加え、匂いの寄与も大きい(図1)。畜肉の匂いの理解は、畜肉の高付加価値化や加工食品の開発につながるなど重要性が増してきている。
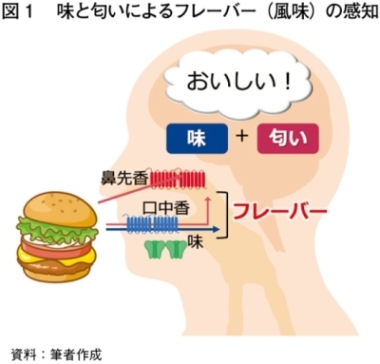
畜肉のフレーバー形成において、加熱は重要である。畜肉を加熱すると、畜肉中のアミノ酸、ペプチド、炭水化物などがメイラード反応(糖とアミノ酸〈タンパク質〉などが結合する反応)や脂質分解反応を引き起こし、加熱した畜肉特有の香ばしいフレーバーが生成される1)。加熱した畜肉に含まれる香気成分として、牛肉ではHexanal、Nonanal、Heptanalなど2)、豚肉ではHexanal、1-Octen-3-ol、(E)-2-Octenalなど3)、鶏肉では2-Furanmethanethiol、2-Methyl-3-Furanthiol、3-Methylthiopropanalなど4)が報告されている。畜種によってフレーバー形成に関与する成分はさまざまである。さらに加熱調理によって生成された多数の香気成分が複合的に寄与し、複雑な加熱畜肉フレーバーが形成される。
牛肉にはさまざまな品種が存在するが、日本人の約7割がおいしさの面で和牛肉を好むことが報告されている5)。和牛肉のおいしさには大きく分けて二つの「香り」が関与している。一つ目は加熱前の生肉で感じる、甘いミルク様の「生牛肉熟成香」であり6)、二つ目は加熱によって生じる甘く脂っぽい「和牛香」である。特に和牛香は和牛肉が好まれる主要な要因になっており、赤身と脂身の共存部に酸素が接触し、加熱時の化学反応により生じると考えられている。和牛香で感じられる甘さには、ココナッツや桃様の香気成分であるラクトン類が関与しており、γ-Decalactone、γ-Undecalactone、δ-Undecalactone、γ-Nonalactone、γ-Hexalactoneなどの成分が報告されている6、7)。ラクトン類の一部は和牛肉の熟成過程で起きるヒドロキシカルボン酸の分子内脱水縮合によって生成されるが、すべてのラクトンの生成経路は完全には明らかとされていない。また、脂っぽさには脂肪様の香りのするDiacetylやAcetoin、一部のアルコール類、アルデヒド類が寄与することも報告されている6)。
(2)ヒトの嗅覚受容体の応答パターンについて
これまで、多くの研究によって畜肉の匂いに関する知見が蓄積されており、高付加価値化や加工食品の開発に応用されてきた。しかし、匂いはヒトの繊細な感覚であり、また曖昧な言葉で表現されるため、官能評価や成分分析などによって客観的に評価することが難しいことは、依然として大きな課題である。一方、近年の分子生命科学の進展により、ヒトは約400種類の嗅覚受容体の応答パターンによって1兆種類以上の匂いを感知・識別できることが明らかとなった(図2)8、9、10、11)。言い換えると、それぞれの匂いに対する嗅覚受容体応答パターンを解明できれば、匂いをデジタルデータとして提示し、客観的・定量的に評価することが可能となる。本研究でわれわれは、ヒト嗅覚受容体応答の網羅的解析システムを開発し、米国産牛肉と黒毛和牛肉をモデルとして、嗅覚受容体の応答パターンを解析した。

2 ヒト嗅覚受容体の応答評価について
(1)ヒト嗅覚受容体の応答評価系
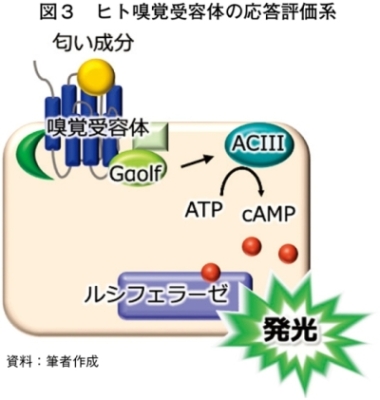
ヒト嗅覚受容体は、HEK細胞などの哺乳類細胞で発現させることで、その応答をin vitro(人為的に作った環境)で評価することが可能である。嗅覚受容体はGタンパク質共役型受容体の一種であり、細胞内シグナルを生じさせるGタンパク質としては、in vivo(生体)と同じGs型のGαolfが用いられることが多い。ヒト嗅覚受容体の活性化によってアデニル酸シクラーゼを介して細胞内でセカンドメッセンジャー(cAMP)が産生され、cAMP濃度依存性の改変型ルシフェラーゼによって受容体応答を発光量として定量的に評価できる(図3)。この際、ヒト嗅覚受容体の細胞表面発現が非常に困難であるため、RTP1Sなどのシャペロン分子を共発現させることで、受容体発現量と応答感度を向上させることが不可欠である。われわれは380種類の全ヒト嗅覚受容体を対象として、最適化した人工遺伝子を合成し、さまざまな匂いに対する受容体応答の評価を進めている。
本研究で用いた細胞は、Flp-InþT-RExþ-293細胞をベースとして、ヒト嗅覚受容体の応答に関わるアクセサリー分子(GαolfやRTP1Sなど)を安定的に共発現させたものである。当該細胞にヒト嗅覚受容体とルシフェラーゼの遺伝子を導入後、384 well plateに播種 、受容体発現細胞にサンプルを作用させ、発光量を測定することで受容体応答を定量解析した。
(2)ろ紙カバー法による気相応答評価系
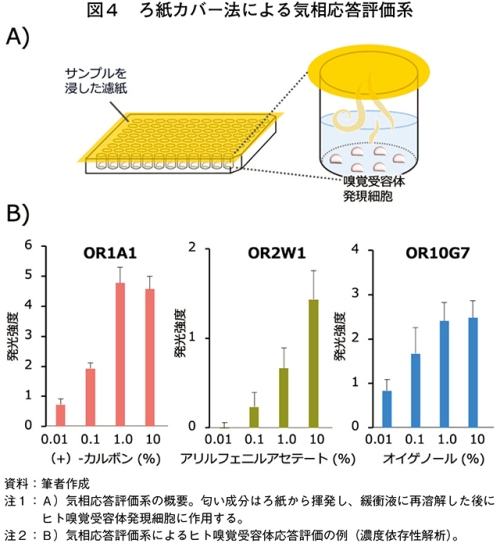
ヒトが食品の匂いを感知する過程では、揮発した匂い成分が嗅粘膜に再溶解された後に嗅覚受容体に作用する。しかし、従来の応答評価系の多くは、ヒト嗅覚受容体発現細胞を浸した緩衝液に液体状のサンプルを投与するものであった。われわれは、ヒト嗅覚受容体発現細胞を播種したプレートを、サンプル溶液を染み込ませたろ紙で覆うことにより、気相を介して効率的に匂い成分を作用させる気相応答評価系を開発した(図4)。本系は従来の液相応答評価系と比較して感度の点では若干劣るものの、各成分の濃度依存的な解析も可能である。また、(1)ヒトの嗅覚知覚システムを模倣した評価が可能(2)揮発成分以外が作用しないため、細胞の非特異的応答が起きにくい(3)固体や気体のサンプルも評価可能(4)ヒト嗅覚受容体発現細胞への個別のサンプル投与が不要(5)成分が混在しているサンプルであっても応答評価できる−などの利点がある。
3 牛肉サンプルの調製
米国産牛肉および黒毛和牛肉(静岡県御前崎市の銘柄牛 遠州夢咲牛)は、赤身と脂身に切り分け、匂いサンプルの調製に用いた。170度に熱したフライパンで脂身を3分間加熱し、油脂を溶解させた後、赤身を加えて4分間加熱した。その後、フライパン上の油脂を浸み込ませたろ紙を、ヒト嗅覚受容体発現細胞を播種したプレートの上に乗せ、30分間、揮発性の香気成分を嗅覚受容体に暴露させた。
4 牛肉香によるヒト嗅覚受容体応答パターンの解析
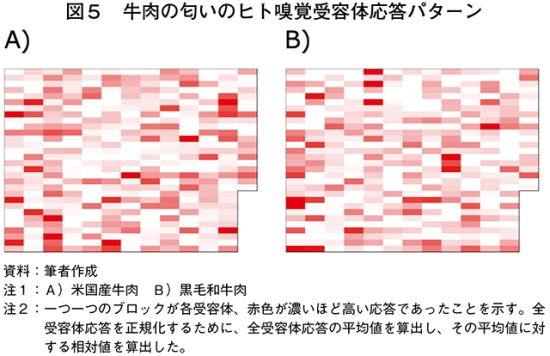
(1)米国産牛肉と黒毛和牛肉の匂いに応答した受容体の比較
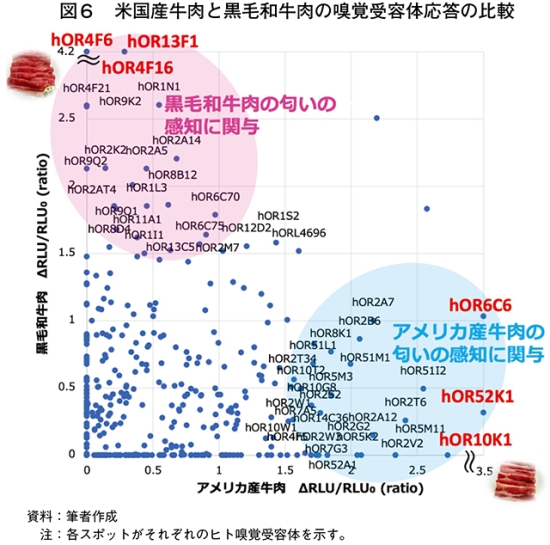
2種類の牛肉香の違いをヒト嗅覚受容体応答パターンとして比較するために、米国産牛肉と黒毛和牛肉をモデルとして、380種類のヒト嗅覚受容体応答を評価した。その結果、米国産牛肉と黒毛和牛肉の匂いは、それぞれ特有のヒト嗅覚受容体応答パターンを示した(図5)。米国産牛肉と黒毛和牛肉の匂いに応答した受容体を比較すると、米国産牛肉で応答値が高く、黒毛和牛肉で応答値が低かった受容体(hOR6C6、hOR52K1、hOR10K1など)は米国産牛肉の匂いの感知に関与している可能性が考えられた(図6)。一方、米国産牛肉で応答値が低く、黒毛和牛肉で応答値が高かった受容体(hOR4F6、hOR4F16、hOR13F1など)は和牛香の感知に関与している可能性が考えられた。このように、言葉では表現しにくい2種類の牛肉の匂いの違いを、ヒト嗅覚受容体応答パターンとして“みえる化”できることが示された。
前述したように、畜肉のフレーバー形成において加熱は重要であり、特に加熱に伴って起きるメイラード反応と脂質の分解がフレーバー形成に大きく寄与している。特に牛肉は他の畜肉よりも脂質が多く含まれるため、調理中に多くの脂質由来の香気成分が生成される。加熱した牛肉の香りに含まれる揮発性成分として、Hexanal、Nonanal、Heptanal、Pentanal、(E,E)-2,4-Decadienalなどが報告されている2)。また、特に黒毛和牛肉は濃厚な甘い「和牛香」が特徴であり、その成分としてγ-hexalactone、γ-decalactone、γ-undecalactoneなどのラクトン類が報告されている7)。それらの成分が、本研究で応答が検出されたヒト嗅覚受容体を活性化させる可能性がある。
畜肉を加熱した際に生成する成分数は1000以上ともされており、当然、前述の成分以外にも多くの成分が匂いの形成に関与していると考えるのが自然である。また、香気成分は単独で知覚されるだけでなく、他の成分との直接的相互作用、あるいは受容体応答パターンに影響する間接的相互作用によっても、複雑な匂いを形成する。本研究で見いだされた応答受容体のパターンを活用することで、それらを統合した、ヒトの感覚に近いデジタルデータとして牛肉の匂いを扱うことができると考えられる。
(2)研究結果の活用と今後の展望について
本研究では、ヒト嗅覚受容体の応答パターンとして、米国産牛肉と黒毛和牛肉の匂いを“みえる化”した。得られた結果は、畜肉の種類や産地、等級などの違いを、受容体応答パターンの違いとして客観的かつ定量的に取り扱うことができることを示唆しており、新たな畜肉の「おいしさ評価系」としての活用が期待できる。一方、それぞれの牛肉の解析で応答受容体として見いだされた受容体について、どの香気成分によって生じた応答なのかは現状では特定できていない。加熱牛肉の香気成分についての報告は数多くなされているものの、匂いは牛肉そのものの種類や部位、脂肪含有量、熟成日数、さらに加熱時の温度や時間などさまざまな条件の影響を受けるため、受容体応答に寄与する成分を特定することは非常に困難である。さらに、成分分析で検出されないほど微量であっても、匂いへの寄与が大きい成分も存在するため、成分ごとに受容体応答を特定するのではなく、牛肉香を包括的に評価、理解することが重要と考えられる。
5 おわりに
本技術により、さまざまな畜肉の匂いを“みえる化”し、それらの違いを客観的かつ定量的に評価することが可能となる。このような方法論は、畜肉のおいしさの理解を深めることなどにおいて役立つと期待できる。ただし、得られた嗅覚受容体応答パターンとヒトの感覚との結び付きは未解明であり、見いだされた応答受容体の活性化によって実際にヒトが「牛肉の匂い」を感じるのかどうかは現時点では不明である。また、成分分析によって特定されている、匂いへの寄与が大きい成分と応答受容体を対応させることや、応答受容体の特定精度の向上(受容体応答感度の向上)、また解析にかかる手間やコストの低減は今後の課題として残されている。畜肉の匂いを客観的かつ定量的に評価できるようになれば、畜産物の差別化を進める際の一つの手法となり、畜産農家の経営安定などにもつながることが期待されるため、実用化に向けたさらなる技術開発が重要である。
参考文献
1. Ba, H., Hwang, I., Jeong, D., Touseef, A. Principle of meat aroma flavors and future prospect. Latest Research into Quality Control Edited by Isin Akyar, 2, 145-176 (2012).
2. Kerth, C., Miller, R. Beef flavor: a review from chemistry to consumer. J. Sci. Food Agric., 95, 2783–2798 (2015).
3. Chang, Y., Wang, S., Chen, H., Zhang, N., Sun, J. Characterization of the key aroma compounds in pork broth by sensory-directed flavor analysis. J. Food Sci., 86, 4932-4945 (2021).
4. Aliani, M., Farmer, L. Precursors of chicken flavor. II. Identification of key flavor precursors using sensory methods. J. Agric. Food Chem., 53, 6455-6462 (2005).
5. 公益財団法人日本食肉消費総合センター 消費動向調査「食肉に関する意識調査」(令和4年10月):http://www.jmi.or.jp/info/survey_files/file0/69.pdf
6. 松石昌典 牛肉の香りと熟成, 日本味と匂学会誌, 11, 137-146 (2004).
7. Ueda, S., Yamanoue, M., Sirai, Y., Iwamoto, E. Exploring the characteristic aroma of beef from Japanese black cattle (Japanese wagyu) via sensory evaluation and gas chromatography-olfactometry. Metabolites, 11, 56 (2021).
8. Malnic, B., Hirono, J., Sato, T., Buck, L. Combinatorial receptor codes for odors. Cell, 96, 713-723 (1999).
9. Bushdid, C., Magnasco, M.O., Vosshall, L.B., Keller, A. Humans can discriminate more than 1 trillion olfactory stimuli. Science, 343, 1370-1372 (2014).
10. Niimura, Y., Matsui, A., Touhara, K. Extreme expansion of the olfactory receptor gene repertoire in African elephants and evolutionary dynamics of orthologous gene groups in 13 placental mammals. Genome Res., 24, 1485-1496 (2014).
11. Buck, L., Axel, R.: A novel multigene family may encode odorant receptors: a molecular basis for odor recognition. Cell, 65, 175-187 (1991).